So far, this pretty morphotype has only been found at two sites, a brown-water lake in Ottawa’s Mer Bleue Bog, and a roadside mire in the Cree Nation of Chisasibi, on the Quebec coast of James Bay.
Our paper will spend 2025 behind a paywall (after which access will be free). In the meantime, a good description of the critter is available on Ferry Siemensma’s site. And here’s some video:
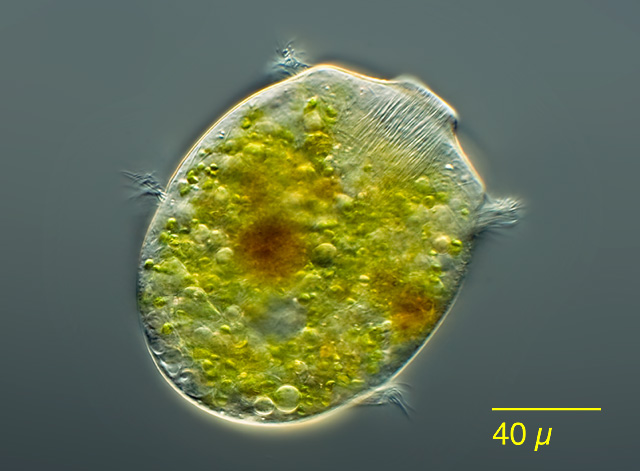
Netzelia corona is a testate amoeba with “teeth” around the aperture of its shell. It also has some impressive spines jutting out from its posterior, which have obvious advantages as a defensive apparatus. But it’s the teeth that have always puzzled me. They’re very neatly made, featuring uniform lobes of organic cement from which tiny, sharp mineral particles protrude.
What is their purpose?
SEM of Netzelia corona, collected in Mer Bleue Bog, Ottawa. Image by Bruce Taylor.
A recent article by Kenneth Dumack and colleagues offers a possible explanation, but it requires a shift in our usual view of the arcellinid shell. Amoeba tests are conventionally seen as protective structures, designed to thwart predators and parasites, and provide shelter in times of drought. However, according to Dumack and his collaborators, the shell is not merely a refuge, but an instrument of predation. As the authors put it in their provocative title: “It’s time to consider the Arcellinida shell as a weapon.”
This insight hinges partly on the behaviour of a protein called “filamentous actin”, a substance that enables amoeboid cells to move forward by extending pseudopods. When a pseudopod forms at the leading edge of the cell, long chains of f-actin molecules rapidly polymerize within it, forming an inner scaffolding of filaments that push the pseudopod out. Then, as the pseudopod is retracted, this scaffolding is quickly disassembled and sections of filamentous actin are pulled back into the body of the cell, with the help of myosin motor proteins.
During locomotion, there is a concentration of f-actin in the pseudopods themselves, which can be seen with the help of a stain made of phalloidin. This substance–originally derived from the Death Cap mushroom, Amanita phalloides–binds to the actin in a cell, and can be combined with fluorescent dyes, so that, in a properly equipped microscope, it takes on a vivid green fluorescence, revealing the exact regions in which actin is concentrated. This was beautifully recorded in an earlier paper (Estermann et al., 2023), using the testate amoeba Cryptodifflugia oviformis. When that amoeba is in motion, we see a high concentration of stained actin in the pseudopods, but nearly none in the body:
Cryptodifflugiaoviformis in locomotion, showing a concentration of green-stained actin in the pseudopods. Source: Estermann et al., 2023
During predation, however, the distribution of actin is quite different. When the same species of amoeba is feasting on fungal hyphae we see filaments of stained actin extending up into the body of the cell.
Cryptodifflugia oviformis preying on strands of fungi. Green stain indicates the presence of actin. Source: Estermann et al., 2023
According to the authors, the actin filaments are anchored on the walls of the shell itself, within the strands of cellular matter traditionally called epipodia (attachment points for the organism, within its test). This is where the shape and structure of the shell–including its aperture–becomes important. In the model proposed by Dumack and associates, as prey is dragged into the shell to be consumed it is broken across the lip of the aperture. Here’s a helpful pair of images, showing the mechanism by which Cryptodifflugiaoviformis breaks open fungal hyphae:
Evidently, the conical bundles of actin shown in the images on the left are used “to exert force on the cell walls of the amoeba’s prey while being anchored to the inner shell surface.” The mechanism of the force is not fully explained at the molecular level, but it seems these actin bundles act somewhat like cables in a winch (presumably driven by contractile proteins, such as myosin?). One end of the actin bundle is firmly attached to the inside of the shell, and the other is affixed to a tasty prey organism. As the victim is pulled into the shell, its membranes or cell walls are broken open on the rim of the aperture.
As for the Netzelia corona I posted, above, I am not sure what organisms it shreds on the spines of its “denticulate lobes”. I’ve never stopped to watch it feed.
However, there is a closely related species, the free-floating carnivorous amoeba Netzelia tuberspinifera, whose shell has a very similar design. This one-celled hunter is capable of taking down large prey, including rotifers, a behaviour that was caught in an interesting sequence of photographs published in a study of “carnivory and active hunting” in that species (Han et al., 2008). The sequence shows N. tuberspinifera penetrating the gelatinous sheath around the rotifer Collotheca, then dragging the animal itself into its shell, to be devoured:
Netzelia tuberspinifera preying on the rotifer Collotheca. Source: Han et al., 2008
The amoeba gains access to the rotifer by breaching the gelatinous sheath that surrounds it (identified in the fourth panel by a dotted line). This does seem quite similar to the feeding strategy described by Dumack et al., and to me it seems likely that the denticulate structures around its aperture play a role in perforating the sheath.
In other words, these “teeth” might actually serve as teeth!
Two wrongs don’t make a right. Fifty wrongs, however, can make a scientific consensus. All it takes is a large enough number of researchers drawing similar conclusions from a big enough pile of bad data. A confirmation cycle sets in, and tentative suggestions begin to solidify into confident assertions. A weird idea that might be true becomes the thing that everybody knows.
Eventually, if the system is working properly, some spoilsport may return to the evidence and go through it all, item by item. If it turns out that the data and methods don’t support previous conclusions, the consensus melts away.
As soon as I saw them, I recognized these “ciliates.” I’d seen the very same thing in my own saliva, almost twenty five years earlier!
A little research explained both cases: these were not protozoans at all, but ordinary ciliated epithelial cells–normal tissues from human lungs and nasal passages. It’s something that happens, occasionally: a few ciliated cells are torn loose, and keep on moving until their energy reserves run out.
Lophomonas blattarum. Source: Beams et al., 1961, adapted from R.R. Kudo
As far as I was concerned, that little mystery was solved.
But while learning about this phenomenon, I ran into something rather creepy: medical case reports of a rare respiratory infection in human patients caused by an organism identified as Lophomonas blattarum.
This was NOT a creature that should be living in anybody’s lungs! Lophomonas blattarum is a parabasalid flagellate, an anaerobic protozoan normally found in the hindguts of cockroaches. Yet there was a growing body of literature describing it an opportunistic human parasite.
At the time, I was skeptical of these case reports, because the microscopy in them was uniformly awful, and the authors made little attempt to reconcile their murky images with the known cellular features of Lophomonas blattarum. The morphological grounds for identifying these “organisms” as Lophomonas struck me as weak, however strong the clinical evidence might be.
Still, without a gene sequence from the supposed pathogen, it seemed like the question could not be put to rest.
Among the comments on my post were some helpful observations by Gillian Gile, who–unlike any of the clinicians who had diagnosed Lophomoniasis in their patients–had actually worked with the organism. At that time, she wrote: “I’ve seen Lophomonas blattarum from cockroaches, and the bundle of flagella moves quite differently from the waving cilia in that youtube video from Mexico. The real L. blattarum can actually swim from point A to point B. With no molecular evidence the jury is still out, but it doesn’t seem likely that Lophomonas is infecting human lungs.” (My emphasis)
Fakhar et al., 2019
It would be a few more years before the first molecular evidence did appear. The results, when they came, were a big surprise to me. In 2019, a group of parasitologists in Iran announced an apparent confirmation, by molecular methods, that Lophomonas was indeed present in a patient with bronchopulmonary disease. They published a paper with a justifiably self-congratulatory title: “First Molecular Diagnosis of Lophomoniasis: the End of a Controversial Story.”
The investigators took a sample from the nasal discharge of a 40 year old woman who was suffering from “rhinorrhea, sneezing, coughing, itchy throat and headache for a month.” They looked at a stained specimen of the discharge under a microscope and found it “positive for Lophomonas.”
A cell identified as Lophomonas blattarum in Farakh et al., 2019.
They included a single image of the supposed organism, and it is as unconvincing as any I’ve seen before–a low-resolution micrograph showing a cell with a broad thatch of cilia distributed evenly across the anterior, rather than a narrowly concentrated apical “horsetail” of flagella, as is typical of Lophomonas.
The authors designed what they called “genus-specific” PCR primers, to selectively amplify a certain chunk of DNA (the SSU rRNA gene, widely used for phylogenetic work). If Lophomonas was present, these primers ought to amplify only the DNA from a certain gene in that particular organism.
They extracted DNA from their patient’s nasal discharge, along with a second sample from a healthy subject as a control, and then performed PCR on both.
Gel electrophoresis confirmed that genetic material from Lophomonas was present in the sick woman’s discharge, and not in the healthy control.
So, it seemed to be true: a gene from a cockroach symbiont really had been found in this poor woman’s lungs! She was treated with an antiprotozoal medication called metronidazole, and she got better. This did look like “the end of a controversial story”, as the paper’s title put it.
Subsequent clinical studies built on this work. New case reports appeared, and in many of them it was now stated as an established fact that Lophomonas blattarum could infect people. Numerous videos confidently labelled “Lophomonas spp.” were posted to YouTube. In 2022, another Iranian study was conducted, and the results were particularly startling. The authors examined 132 frozen bronchial lavage samples from patients hospitalized with a variety of respiratory conditions. Using similar methods to those described in Fakhar et al. (2019), they found that more than 1/4 of these patients (27.3%) tested positive for Lophomonas! A condition previously understood to be rare, was now described as “a common and emerging disease in the study area, southwestern Iran.”
Nguyen et al., 2023
Lophomonas blattarum, from Nguyen et al., 2023. The vesicles labelled “y” are yeast cells. The scale bar is 20 μm.
The authors of that study dissected some actual cockroaches, and succeeded in establishing cultures of Lophomonas blattarum. They produced good photographic images of them, finally, and the images corresponded very well with illustrations in previous studies (like the one from Beams et al., posted above), showing a very narrow tuft of flagella at the apex of the cell, and a nucleus in the anterior of the cell. The organism in their images did not look much like the photos and videos of the supposed “Lophomonas” found in human “infections”.
The authors sequenced these cockroach commensals, and established their phylogenetic position, branching close to a genus of parabasalid termite symbionts called Trichonympha. They also looked more closely at the earlier sequences of putative “L. blattarum” taken from human samples, and found they were not closely related to Lophomonas blattarum at all, but were actually 99% similar with certain members of a different group of organisms called Trichomonadida (two genera in particular, Tetratrichomonas and Pentatrichomonas, one of which is known to live inside human bodies, and looks nothing like either Lophomonas or a ciliated epithelial cell).
A cell found in a patient’s sputum, wrongly identified by Iranian clinicians as “Lophomonas blattarum” (Nasseri et al., 2022) Note the broad carpet of cilia covering the apex of the cell, and compare the distribution of flagella in the image from Nguyen et al., 2023, above.
As it turned out, none of the sequences taken from sick people in previous studies actually belonged to Lophomonas: “These data…indicate that no true L. blattarum sequences have yet been published from human lung samples.” (Nguyen et al., 2023)
The results are pretty devastating for this “emerging human pathogen”.
The authors analyzed all the photographic and video evidence in the published literature, applying the known morphological criteria for identifying Lophomonas. They found that none of the existing images of Lophomonas from human samples showed the characteristic features of the organism, such as a “tight anterior bundle of flagella, an anterior nucleus, the calyx surrounding the nucleus, or a posteriorly protruding axostyle.”
As it turns out, the differences between true Lophomonas and human epithelial cells are not subtle at all. Consider the following figure. The first three panels (A-C) show Lophomonas blattarum, and the second three (D-F) show human epithelial cells. Note the shape of the flagellar bundle in the anterior, the location of the nuclei (labelled “n”), and the overall appearance of the cell.
Source: Mewara et al., 2024
The video evidence is even more clear. The supplementary materials to the article include footage of genuine Lophomonasblattarum which can be downloaded and viewed. In that video, taken by Gillian Gile, a Lophomonas cell is seen moving purposefully forward, with the help of a narrowly concentrated tuft of flagella at the apex of the cell. Distinctive features of the cell, such as the axostyle and the calyx (both recorded by R. R. Kudo in the 1920s!), can be seen at certain points.
Source: Mewara et al., 2024. Used with permission.
The efficient, directed movement of the cell has little resemblance to the feeble, rocking motion we often see in video of “Lophomonas” taken from human samples. Here is a typical example of that:
Source: Peeyush1980 channel on YouTube
In another video from a recent paper, we see a cell with a broadly distributed “carpet” of cilia, going around in circles. This one is more vigorous, but its movements are just as aimless and ineffectual:
In their review of the literature, Mewara et al., 2024 also address the molecular evidence, reaffirming what had been shown in Nguyen et al.,2023: that the PCR assay used to amplify Lophomonas from BALF was not genus-specific at all, and readily amplified other organisms (in particular, certain trichomonads already known to live in human lungs and GI tracts).
Finally, the authors reviewed all the existing clinical evidence for “lophomoniasis,” and the medical case for the ailment seems to have fared just as badly. They found little consistency in the presentation of the “disease”, the ages or backgrounds of those affected, or the actual site of the infection. Evidence needed to single out L. blattarum as the causative agent was missing from the work that had been done.
Their meta-analysis uses methods and standards I don’t really understand, and I’m not competent to redescribe their results. Luckily, I don’t have to, because two of the authors have discussed their work in a conversation with two editors from the Journal of Clinical Microbiology. The video of that conversation has been posted to YouTube. If you’ve never considered the possibility that your lungs could be colonized by creatures from a roach’s rump, you probably don’t need to watch the video. However, if, like me, you’ve already been “infected” by this horrible idea, you might find it comforting:
When conditions for life are unfavourable, some protists can cheat death by entering a cryptobiotic state. Usually, they do it by enclosing themselves within thick-walled structures called “resting cysts,” which protect vital genetic material from dehydration and extreme temperatures. Tucked away in their cysts, the cells shut down all metabolic activities and wait for the environment to improve.
Meanwhile, up in the higher branches of the tree, molecular phylogenetics continues to shuffle things around. My last post (dated November, 2016!) discussed a novel amoeboid called Lecythium siemensmai. That species is no longer a Lecythium at all, but a member of a genus called Fisculla (named for its resemblance to a bag of money). I should have updated that post seven years ago!
My happy place
A few things have changed in my life, too. I’m still fascinated by ciliates, but have been spending most of my microscope hours on testate amoebae. This shift in focus was prompted, in part, by some new tools that have been made available to me.
I now have a small perch at the Canadian Museum of Nature, as a Research Associate. The title comes with a keycard that opens doors at the Natural Heritage Campus in Gatineau, where our country keeps its mastodon bones, its bottled molluscs, and its collection of over a million beetles. This magic card also gives me access to some nice microscopes. The most exciting of these, for me, is the museum’s FEI Apreo II Scanning Electron Microscope, which can resolve features that are measured in nanometres (that is, billionths of a metre!).
Here, for example, is a creature I collected in Ottawa’s Mer Bleue Bog, a snail-shaped testate amoeba called Lesquereusia spiralis, which assembles its shell from curved siliceous rods (which remind me of Cheez Doodles™):
Similar meshes are seen in other lobose shelled amoebae (members of the order Arcellinida), but they often differ in appearance. Back in the 1980s, some researchers proposed that different arrangements of organic cement might be used to distinguish taxa within the group, but the idea doesn’t seem to have caught on. The structure of the organic cement is sometimes noted in species descriptions, but I’m not aware of any attempt to use it a taxonomic character for genus or family. Do patterns in the organic cement correspond with lineages of arcellinids, above the level of species? I have no idea, but it seems like a fun thing to wonder about.
There’s a natural pond down the road, where I collect samples. I always go to the same spot, the edge of a shallow basin in the “littoral zone”, where the pond shades into the shore. I follow the same routine each time: I dip a turkey baster into the soft muck–organic sediments enriched with duckshit and decomposing leaves–and suck out a few cc’s for the bottom of the jar; then I ladle a bit of surface water over the mud, to create a miniature approximation of the natural site.
Sampling Site in Turtle Pond
I’ve taken hundreds of samples there over the years, from an area no bigger than a coffee table. And, despite having done this regularly for about half a decade, nearly every jar I bring home contains something I’ve never seen before.
Of course, to a microbe, a coffee table-sized site is actually quite big. If you were a typical one-celled protist–say, 50 micrometres from tip to tail–a four-foot patch of pond would be more than 24,000 times longer than your body. From a microbe’s point of view, then, my sampling basin is equivalent to a lake 28 miles long. Scaled up to human size, the entire pond would be at least 1,350 miles across, with a greater surface area than the Mediterranean Sea! So, there’s quite a lot of available habitat in the small body of water I call “Turtle Pond.”
At any given time, only a small fraction of the site’s potential diversity will be visible. The mix of species can change from one hour to the next, as weather and random events alter water chemistry, light levels, temperature and organic nutrients. A few hours of sunlight might kick off a sudden bloom of cyanobacteria, soon followed by nassulid ciliates that eat strands of blue algae like spaghetti; then, a few minutes of rain disperse the bloom, the nassulids disappear, and are replaced by some other combination of species. The new populations have barely established themselves, when a turtle (which, from a microbe’s point of view, is about 7 miles long), blunders into the shallows, churning up mud, mixing oxygen-rich upper layers with the anoxic sediments below, and everything changes again. Each shift in the pond creates new opportunities for some organisms, and makes life impossible for others. One species blooms, and another goes into dormancy, withdrawing into resting cysts or simply lurking in reduced numbers, waiting for better days.
The microbial “seed bank” can store a staggering diversity of such hidden opportunists, which emerge from their cysts only when conditions are exactly right. Some are “weed” species, generalists that crop up in nearly every sample. Others have very particular needs, and rarely appear. A few may never find what they need to grow and reproduce. If any cysts of salt-loving (halophilic) protists should blow in from some distant shore, they are probably out of luck.
Recently, I brought home a few jars from the usual spot, and found three rather flamboyant species to cross of my life list. One was Actinobolina radians, an oddball I’ve been hoping to find for years (ever since I first saw it’s mugshot in Vladimir Schewiakoff’s beautifully-illustrated Infusoria Aspirotricha of 1889). Actinobolina is a ciliate in the class Litostomatea, a group of rapacious and morphologically hilarious carnivores. The class includes such celebrity ciliates as the improbably elastic Lacrymaria, the elephant-nosed Dileptus, and a frisky little keg full of death called Didinium, which eats other ciliates like popcorn. Like its cousins, Actinobolina is a hunter. However, it has a unique strategy for capturing prey. When swimming about, it looks much like any other ciliate: egg-shaped, and uniformly covered with cilia. Once it has a found a suitable feeding site, it comes to a stop and slowly extends a large number of long, stiff tentacles, each of which carries toxic extrusomes (cellular darts) to be released on contact into its victim’s body. In this state, it looks rather like a heliozoan “sun animalcule”. With its tentacles extended, Actinobolina lies in wait like a spider in its web, until some tasty creature–a rotifer , or another ciliate–bumps into it. The toxic extrusomes are deployed and the victim is paralyzed, whereupon the ciliate withdraws its tentacles, pulling the stunned prey toward its cellular mouth.
Unfortunately, I didn’t manage to record a successful hunt, but I did get some footage of the ciliate extending and retracting its tentacles. And really, I was just pleased to have seen this bizarre organism.
Another creature that appeared in my jars was the odd, oxygen-hating amoebozoan Pelomyxa, an indiscriminately hungry thing that looks like a tumbling bag full of rocks and food. I see Pelomyxa fairly often, but this one was a different species, smaller and more colorful, and with a distinctive style of movement, featuring frequent lateral “eruptions” of pseudopods. I believe it is Pelomyxa binucleata, a species first identified in the 19th century. For a time, in the late 20th century, it was believed that P. binucleata and all “species” of Pelomyxa, were really just forms and phases of a single species, Pelomyxa palustris. However, since 2004 the Russian researcher Alexander O. Frolov and his collaborators have been repopulating the genus, discovering new species and rehabilitating older ones. There are more than half a dozen, now.
Pelomyxa is also interesting because of its role in the rise and fall of the fabled Lost Protist Kingdom, once known as Archezoa. At one time, in the 1980s and early 90s, Pelomyxa was believed to be a primitive eukaryote, a “living relic of the Proterozoic era” (Margulis & Sagan, 1990) lacking such basic organelles as mitochondria, centrioles, flagella/cilia, endoplasmic reticulum, Golgi bodies and even chromosomes (Margulis & Chapman, Kingdoms and Domains, 2009). It was seen as sort of a missing link between eukaryotes and their bacterial predecessors, and was placed along with an assortment of other anaerobes, in the short-lived Kingdom Archezoa. However, that hypothesis began unraveling within just a few years. Pelomyxa were found to have centrioles and flagella after all, as well as mitochondrial remnants (“mitochondria-derived organelles”). In fact, they turned out to possess most of the stuff you’d expect to find in a eukaryotic cell, along with some nifty new adaptations to anaerobic living, so there was really no reason to regard them as “primitive” at all. When genetic analysis of the genus was finally done, it was placed squarely within the phylum Amoebozoa. Archezoa itself–briefly a great Kingdom, ruled by a benign despot named Thomas Cavalier-Smith–was quietly removed to the museum of obsolete evolutionary hypotheses.
Finally, my samples turned up a single specimen of Supraspathidium, a rather long haptorid ciliate with wide, blubbery lips. I’m afraid this guy will be of no interest to anyone but the most devout ciliatophile, but I’m posting it anyway because it’s my blog. Supraspathidium resembles ciliates of the better-known genus Spathidium, except that it has numerous contractile vacuoles, instead of a single big one in the posterior of the cell. This one has a long, wormy macronucleus, and generally resembles a spathidiid described by Eugène Penard in 1922,under the name Spathidium vermiforme.
In my previous post on the lorica-dwelling scuticociliate Calyptotricha pleuronemoides, I mentioned that only two substantial articles have been written about the species since its discovery (apart from brief descriptions in various places). Until yesterday, I’d been unable to find the second article, which appeared in the German microscopy journal Mikrokosmos in 1999. Luckily, one of the co-authors, Martin Kreutz, was kind of enough to send me a copy!
Calyptotricha pleuronemoides. Image by Martin Kreutz. Source: micro*scope. Click on image for link to source.
Martin tells me he sees the organism frequently in water from the sphagnum ponds of Simmelried, a system of bog lakes, like the one in Ottawa’s Mer Bleue where I sometimes find Calyptotricha. He and Philipp Mayer provide a good redescription of the ciliate, with morphometrics. Their measurements match those of the specimens I’ve found, and agree with those of Phillips and D.S. Kellicott. (All sources give a size range somewhat smaller than that recorded by Alfred Kahl, who gives 50 µm for the length of the cell, and 85 µm for the lorica).
The Mikrokosmos article is difficult to find, so I thought I might give a brief redescription of the species, based on the information collected by Kreutz and Mayer:
Calyptotricha pleuronematoides: Pleuronematid ciliates, in spindle-shaped hyaline lorica 56-75 µm long, tubular with narrowed openings at either end, 9-13 µm in width. Lorica broadens in the middle to a width of 23-24 µm. Cell body resembling Cyclidium, slightly flattened back to front, 22-35 µm long, 15-24 µm wide, somewhat concave on the ventral surface, where oral apparatus occupies 3/4 of body length. L-shaped undulating membrane, made up of fused cilia, 14 µm long. Caudal cilium 10-12 µm long; roughly 17 somatic kineties, spaced 2-3 µm apart. Most specimens with 8-15 zoochlorellae, colorless examples rare. Single oval macronucleus, with small spherical micronucleus. CV in posterior.
The authors mention that specimens are seldom seen outside of their loricas. Free-swimming individuals move quickly, but not as jerkily as Cyclidium. I happen to have recorded a free-swimming Calyptotricha, last year, so I might as well post it here:
References:
Kreutz, Martin, and Philipp Mayer. “Artikel-Calyptotricha pleuronemoides-Ein Ciliat in einer Rohre.” Mikrokosmos 88.1 (1999): 27-30.



















{kind=link}
{kind=link}
{kind=link}
{kind=link}