When conditions for life are unfavourable, some protists can cheat death by entering a cryptobiotic state. Usually, they do it by enclosing themselves within thick-walled structures called “resting cysts,” which protect vital genetic material from dehydration and extreme temperatures. Tucked away in their cysts, the cells shut down all metabolic activities and wait for the environment to improve.
Meanwhile, up in the higher branches of the tree, molecular phylogenetics continues to shuffle things around. My last post (dated November, 2016!) discussed a novel amoeboid called Lecythium siemensmai. That species is no longer a Lecythium at all, but a member of a genus called Fisculla (named for its resemblance to a bag of money). I should have updated that post seven years ago!
My happy place
A few things have changed in my life, too. I’m still fascinated by ciliates, but have been spending most of my microscope hours on testate amoebae. This shift in focus was prompted, in part, by some new tools that have been made available to me.
I now have a small perch at the Canadian Museum of Nature, as a Research Associate. The title comes with a keycard that opens doors at the Natural Heritage Campus in Gatineau, where our country keeps its mastodon bones, its bottled molluscs, and its collection of over a million beetles. This magic card also gives me access to some nice microscopes. The most exciting of these, for me, is the museum’s FEI Apreo II Scanning Electron Microscope, which can resolve features that are measured in nanometres (that is, billionths of a metre!).
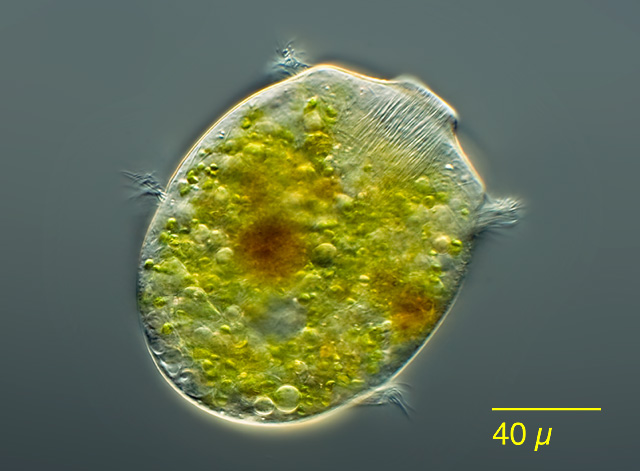
Here, for example, is a creature I collected in Ottawa’s Mer Bleue Bog, a snail-shaped testate amoeba called Lesquereusia spiralis, which assembles its shell from curved siliceous rods (which remind me of Cheez Doodles™):
Similar meshes are seen in other lobose shelled amoebae (members of the order Arcellinida), but they often differ in appearance. Back in the 1980s, some researchers proposed that different arrangements of organic cement might be used to distinguish taxa within the group, but the idea doesn’t seem to have caught on. The structure of the organic cement is sometimes noted in species descriptions, but I’m not aware of any attempt to use it a taxonomic character for genus or family. Do patterns in the organic cement correspond with lineages of arcellinids, above the level of species? I have no idea, but it seems like a fun thing to wonder about.
After studying amoeboid organisms for several decades–observing them in the field, publishing articles about them, and developing his wonderfully detailed website devoted to the little shapeshifters—Ferry Siemensma now has an amoeba he can call his own. The organism, discovered by researchers in a small greenhouse pond at the university of Cologne, is Lecythium siemensmai, a cercozoan amoeboid that lives inside a flexible, secreted transparent “theca”. It looks like this:
The new species, is described, along with several others, in a complete revision of the genus Lecythium, recently published by Kenneth Dumack and two colleagues at Cologne. The genus itself is fairly old—it was erected by Richard Hertwig and E. Lesser in 1874—but it has been a very long time since taxonomists have paid any attention to its composition. Many organisms that properly belong in the group have been misplaced in other genera, such as Gromia, Trinema and Pamphagus. The latter has been recognized since 1915 as an invalid group (the name properly belongs to a genus of grasshoppers), but it is a mark of the neglect into which amoeboid taxonomy has fallen that a Google search on “pamphagus amoeba’’ turns up dozens of modern micrographs and videos of cells identified as belonging to a group that was formally abandoned over a hundred years ago. It is a tribute to the great amoebologists of the past that their work still casts such a long shadow. It is also a reminder that revisions like this one by Dumack et al. are urgently needed.
The paper, charmingly titled “A Bowl with Marbles” (a reference, presumably, to the glassy, round shape of the organism’s soft theca), uses molecular phylogeny to establish the genus as a well-defined clade within Tectofilosida (a rhizarian group whose organic shells lack the siliceous scales secreted by better-known filose amoebae like Euglypha and Assulina). In the section on taxonomy, the authors deftly untangle a few historic hairballs; for instance, tracing the relationship of their Lecythium spinosum to antecedents like Trinema spinosum, Plagiophrys armatus and Pamphagus armatus. Finally, they provide a visual key to the group, featuring clear illustrations and helpful diagnostic clues. For this genus, at least, they have built a convenient foot-bridge between phylogeny and field identification.
Congratulations to Ferry, and to the researchers at Cologne, all of whom are doing the kind of work that could coax future researchers back to the field.
REFERENCES:
• Dumack, Kenneth, Christina Baumann, and Michael Bonkowski. “A Bowl with Marbles: Revision of the Thecate Amoeba Genus Lecythium (Chlamydophryidae, Tectofilosida, Cercozoa, Rhizaria) Including a Description of Four New Species and an Identification Key.” Protist 167.5 (2016): 440-459.
• Hertwig, Richard, and E. Lesser. “Ueber Rhizopoden und denselben nahestehende Organismen.” Arch. Mikr. Anat 10.1874 (1874): 35-243.
There’s a natural pond down the road, where I collect samples. I always go to the same spot, the edge of a shallow basin in the “littoral zone”, where the pond shades into the shore. I follow the same routine each time: I dip a turkey baster into the soft muck–organic sediments enriched with duckshit and decomposing leaves–and suck out a few cc’s for the bottom of the jar; then I ladle a bit of surface water over the mud, to create a miniature approximation of the natural site.
Sampling Site in Turtle Pond
I’ve taken hundreds of samples there over the years, from an area no bigger than a coffee table. And, despite having done this regularly for about half a decade, nearly every jar I bring home contains something I’ve never seen before.
Of course, to a microbe, a coffee table-sized site is actually quite big. If you were a typical one-celled protist–say, 50 micrometres from tip to tail–a four-foot patch of pond would be more than 24,000 times longer than your body. From a microbe’s point of view, then, my sampling basin is equivalent to a lake 28 miles long. Scaled up to human size, the entire pond would be at least 1,350 miles across, with a greater surface area than the Mediterranean Sea! So, there’s quite a lot of available habitat in the small body of water I call “Turtle Pond.”
At any given time, only a small fraction of the site’s potential diversity will be visible. The mix of species can change from one hour to the next, as weather and random events alter water chemistry, light levels, temperature and organic nutrients. A few hours of sunlight might kick off a sudden bloom of cyanobacteria, soon followed by nassulid ciliates that eat strands of blue algae like spaghetti; then, a few minutes of rain disperse the bloom, the nassulids disappear, and are replaced by some other combination of species. The new populations have barely established themselves, when a turtle (which, from a microbe’s point of view, is about 7 miles long), blunders into the shallows, churning up mud, mixing oxygen-rich upper layers with the anoxic sediments below, and everything changes again. Each shift in the pond creates new opportunities for some organisms, and makes life impossible for others. One species blooms, and another goes into dormancy, withdrawing into resting cysts or simply lurking in reduced numbers, waiting for better days.
The microbial “seed bank” can store a staggering diversity of such hidden opportunists, which emerge from their cysts only when conditions are exactly right. Some are “weed” species, generalists that crop up in nearly every sample. Others have very particular needs, and rarely appear. A few may never find what they need to grow and reproduce. If any cysts of salt-loving (halophilic) protists should blow in from some distant shore, they are probably out of luck.
Recently, I brought home a few jars from the usual spot, and found three rather flamboyant species to cross of my life list. One was Actinobolina radians, an oddball I’ve been hoping to find for years (ever since I first saw it’s mugshot in Vladimir Schewiakoff’s beautifully-illustrated Infusoria Aspirotricha of 1889). Actinobolina is a ciliate in the class Litostomatea, a group of rapacious and morphologically hilarious carnivores. The class includes such celebrity ciliates as the improbably elastic Lacrymaria, the elephant-nosed Dileptus, and a frisky little keg full of death called Didinium, which eats other ciliates like popcorn. Like its cousins, Actinobolina is a hunter. However, it has a unique strategy for capturing prey. When swimming about, it looks much like any other ciliate: egg-shaped, and uniformly covered with cilia. Once it has a found a suitable feeding site, it comes to a stop and slowly extends a large number of long, stiff tentacles, each of which carries toxic extrusomes (cellular darts) to be released on contact into its victim’s body. In this state, it looks rather like a heliozoan “sun animalcule”. With its tentacles extended, Actinobolina lies in wait like a spider in its web, until some tasty creature–a rotifer , or another ciliate–bumps into it. The toxic extrusomes are deployed and the victim is paralyzed, whereupon the ciliate withdraws its tentacles, pulling the stunned prey toward its cellular mouth.
Unfortunately, I didn’t manage to record a successful hunt, but I did get some footage of the ciliate extending and retracting its tentacles. And really, I was just pleased to have seen this bizarre organism.
Another creature that appeared in my jars was the odd, oxygen-hating amoebozoan Pelomyxa, an indiscriminately hungry thing that looks like a tumbling bag full of rocks and food. I see Pelomyxa fairly often, but this one was a different species, smaller and more colorful, and with a distinctive style of movement, featuring frequent lateral “eruptions” of pseudopods. I believe it is Pelomyxa binucleata, a species first identified in the 19th century. For a time, in the late 20th century, it was believed that P. binucleata and all “species” of Pelomyxa, were really just forms and phases of a single species, Pelomyxa palustris. However, since 2004 the Russian researcher Alexander O. Frolov and his collaborators have been repopulating the genus, discovering new species and rehabilitating older ones. There are more than half a dozen, now.
Pelomyxa is also interesting because of its role in the rise and fall of the fabled Lost Protist Kingdom, once known as Archezoa. At one time, in the 1980s and early 90s, Pelomyxa was believed to be a primitive eukaryote, a “living relic of the Proterozoic era” (Margulis & Sagan, 1990) lacking such basic organelles as mitochondria, centrioles, flagella/cilia, endoplasmic reticulum, Golgi bodies and even chromosomes (Margulis & Chapman, Kingdoms and Domains, 2009). It was seen as sort of a missing link between eukaryotes and their bacterial predecessors, and was placed along with an assortment of other anaerobes, in the short-lived Kingdom Archezoa. However, that hypothesis began unraveling within just a few years. Pelomyxa were found to have centrioles and flagella after all, as well as mitochondrial remnants (“mitochondria-derived organelles”). In fact, they turned out to possess most of the stuff you’d expect to find in a eukaryotic cell, along with some nifty new adaptations to anaerobic living, so there was really no reason to regard them as “primitive” at all. When genetic analysis of the genus was finally done, it was placed squarely within the phylum Amoebozoa. Archezoa itself–briefly a great Kingdom, ruled by a benign despot named Thomas Cavalier-Smith–was quietly removed to the museum of obsolete evolutionary hypotheses.
Finally, my samples turned up a single specimen of Supraspathidium, a rather long haptorid ciliate with wide, blubbery lips. I’m afraid this guy will be of no interest to anyone but the most devout ciliatophile, but I’m posting it anyway because it’s my blog. Supraspathidium resembles ciliates of the better-known genus Spathidium, except that it has numerous contractile vacuoles, instead of a single big one in the posterior of the cell. This one has a long, wormy macronucleus, and generally resembles a spathidiid described by Eugène Penard in 1922,under the name Spathidium vermiforme.
An elusive giant has risen from the muck at the bottom of a Florida canal. It is the ciliate Loxodes rex, a large and shapely thing once thought to be endemic to equatorial Africa. Hunter Hines, a Ph.D. student affiliated with the Harbor Branch Oceanographic Institution, has published the first record of the species on the North American continent, and has also posted video of the creature on YouTube:
It is not the first time the species has turned up far from its supposed home. Specimens of the big guy have previously been reported from an artificial pond Thailand. It also turns up on a diversity checklist from São Paulo (though identifications in lists of that kind can’t be independently corroborated). The new study provides better images and metrics, although silver staining and gene sequencing were not done.
Its presence in Florida is interesting, because Loxodes rex is one of the 52 ciliate “flagship species” listed by Wilhelm Foissner as good candidates for testing the hypothesis that some species of ciliate have a restricted geographical range (Foissner, 2008). The ciliates on Foissner’s list are all eye-catching creatures easily seen in the light microscope, difficult to misidentify and unlikely to be overlooked in diversity studies. All of them seem to show some evidence of endemism: that is, they have not been found everywhere in the world, but have turned up only in certain regions. This makes them ideal for proving or disproving the hypothesis that ciliates, like orchids and elephants, have biogeography. As Foissner and his collaborators put it: “[T]he hypothesis of restricted distribution of certain ciliate species must be refused when a considerable number of them is found in all or most biogeographic regions.” (Foissner et al, 2008)
Loxodes rex, from Hines et al, 2016
To some readers, it might seem odd that there is any controversy about this. Cave snakes and capybaras have biogeography. Why not ciliates? That question takes us back to a hundred-year-old conjecture by two Dutch biologists, Martinus Beijerinck and Baas Becking, concerning the apparent ubiquity of microbial species. Their speculation was that microorganisms, by virtue of their small size and certain special talents (such as the ability to form resting cysts), were not confined by geographical barriers. Every species could exist anywhere in the world, as long as its preferred environmental conditions are available. If an organism is small enough to waft about in the wind, or travel from place to place in the soggy tailfeathers of a bird, its distribution cannot be limited to any particular region. Baas Becking bundled this idea into a phrase so concise and quotable that it has become almost obligatory to repeat it every time the subject of microbial endemism is raised: “Everything is everywhere, but the environment selects.”
This is sometimes dignified as the “Baas Becking hypothesis,” but Foissner argues that is not a proper hypothesis at all, because it is unfalsifiable (Foissner, 2006). After all, if an organism in one pond fails to appear in another, the difference can always be explained by some small dissimilarity in the local conditions. No two ponds provide identical environmental conditions–indeed, there can be an impressive diversity of micro-environments within a single pond, and local conditions can change from one hour to the next. To test the “ubiquity hypothesis” as expressed in Becking’s memorable phrase, you’d need to have two truly identical bodies of water in two distinct biogeographical regions. It is an impractical experiment, because there are simply too many variables.
The main competitor to “everything is everywhere” is the “moderate endemicity model” advanced by Foissner and others, which holds that while as many as 2/3 of microbial species may be cosmopolitan, the rest are found in only certain places. In other words: Some things are not everywhere. This hypothesis is testable, thanks to the terms Foissner himself has set: if a “considerable number” of the flagship species turn out to be cosmopolitan, moderate endemicity is falsified.
By that standard, it does seem that the newly expanded range of Loxodes rex slightly improves the larger case against ciliate endemism–although, it should be said, Hines does not address Foissner’s stipulation that the flagship species must be found in a site that is not “prone to be contaminated by invaders” (Foissner, 2006). Obviously, the loss of one purported endemic does not constitute a “considerable number,” but the authors of the Loxodes paper also claim to have found “other large ciliates with alleged restricted distributions,” and have already posted video of one, the fairly cool-looking Frontonia vesiculosa. It will be interesting to see what else they turn up.
Dragesco’s drawing of Loxodes rex (A) next to Loxodes magnus (B). Magnus is big, but rex is the King.
While Hines et al. do not show stained specimens, the imaging is fairly clear, and the data provided do seem to show that members of the Florida population of Loxodes rex are morphologically close to the ones Jean Dragesco found in Africa. This this can be explained by dispersal, either through natural means (everything is everywhere) or by recent human activity.
Or, it could also be (and what’s a blog for, if not wild speculation?) that both populations are remnants from a single group that split when the Gondwanan parts of the supercontinent Pangaea broke up some 180 million years ago (Florida, unlike most of North America, was largely attached to Gondwana). If that is true, then phylogenetic analysis of the Florida and Africa populations–if anyone ever gets around to it–would show a lot of genetic divergence (a few hundred millions years’ worth), even though morphology of the species has been conserved. Morphological stasis of a species, even over long stretches of geological time, is unexceptional, as Stephen Jay Gould argued at exhausting length in The Structure of Evolutionary Theory, and recent work by Heger et al. on the testate amoeba Hyalosphenia papilio has shown how much genetic diversity can be concealed behind a highly conserved morphology.
If the Florida population of Loxodes rex turns out to be a stable, genetically distinct subgroup, like the cryptic species Hyalosphenia papilio studied by Heger et al., the notion that ciliates have biogeography remains somewhat intact, while the practice of defining species by small numbers of morphological characters suffers another small blow.
Of course, that moves the goal posts for testing the “ubiquity hypothesis,” but I do feel we need to ask whether the ongoing dispute about whether “everything is everywhere” is, in part, an artifact created by simplistic species concepts.
I’ve finally identified a thing I found in my mouth nearly 25 years ago. Unfortunately, solving that old puzzle has left me with a new one concerning a protozoan normally found in the hindgut of cockroaches and termites.
I ran across it in the fall of 1990. My wife was a first-year medical student and had borrowed a microscope to study samples of human tissue for a course in histology. In those days, I was more interested in poetry than protozoa, but I was even more interested in procrastination. So, one morning I put my doctoral thesis aside and took the loaner ‘scope for a spin.
Her prepared histology slides–fixed cells stained in lurid pinks and powder blues–did not hold my attention for long. I wanted to see something alive and wriggling. I’d read about dust mites, so I put some floor-fluff on a slide, but that was disappointing: nothing but hair, skin flakes and coloured cloth fibres. Then, I sampled some water from a soup pot that had sat in the sink overnight, which was a little better. There were bacteria there, thousands of them, jostling and jiggling in the milky broth. But bacteria are boring.
A reconstruction of The Thing. (The arrows are supposed to show how it moved. Yes, it looks like the cell has arms, but I don’t feel like drawing it again).
Finally, I swabbed some saliva from the back of my mouth and that was more like it. If you’ve ever done this, you’ll know what I found: big, ragged oral epithelial cells, swirling rafts of intriguing organic stuff, plump leukocytes that had recently eaten a bacterial meal. Then something truly bizarre floated into view. It was round and mottled, like a degenerate happy face with a crop of lank hairs sprouting from one side. It was rotating back and forth with a hypnotic, rocking motion: clockwise, then counterclockwise, then clockwise again. With each change of direction, the strands moved together in a languid arc.
Amazing! A parasite was living in my mouth, and it had hair like a rock star. What could it be? I grabbed a bus down to the McGill Life Sciences Library and pulled some parasitology texts from the stacks. From these, I determined that this creature had to be some kind of “flagellate” (the word was new to me). I leafed through the mug shots, blurred micrographs of Trichomonas, Chilomastix, Giardia, Retortamonas and all their troublesome colleagues, but I couldn’t find my guy anywhere. The known flagellate parasites were all were too pinched, or too pointy, and none had the lush head of hair I’d seen on my creature.
A couple of nights later, a burglar broke into our place and stole the microscope, which put an end to my amateur parasitology for a couple of decades. I went back to my real work, but never forgot that my mouth had miniature glam rockers living inside it. Years later, when I returned to microscopy (taking procrastination to a whole new level), the first thing I did was try to find those flagellates again. I swabbed my palate, scraped my tongue and spat on slide after slide, without success. By then, I had learned a lot more about protists, and I had acquired a good library of field guides and protozoology texts, none of which recorded anything quite like the organism I’d seen. I began to doubt my memory.
Then, just two days ago, a friend from Uruguay sent me a link to a video from Mexico, showing a presumed protozoan found in the bronchial lavage of a patient who had nearly drowned in a river. And there it was again, the Thing from my Mouth!
That rocking motion is exactly what I had seen in 1990. The video was posted to YouTube as an unknown “ciliado” (ciliate), and superficially it does resemble one. At first, it struck me as an overripe oligotrich, possibly bloated from osmotic pressure and moving poorly. But the way it was moving looked all wrong, even for an unhealthy specimen. That repetitive, ineffectual rocking movement was not like the familiar motion of the fused cilia (adoral polykinetids) that surround the cellular mouth of oligotrich ciliates, or any of the ciliary wreaths and fringes I knew from other free-living ciliophora. The rocking suggested something sessile that had come loose from its substrate, which reminded me that this “ciliate” had been found in bronchealveolar lavage fluid (BALF). As it happens, we all have ciliated cells lining some of our nasal and bronchial cavities, which perform the useful task of clearing mucus and debris from the lungs. The ciliated cells stay fixed in one place, and waste matter sort of “crowd surfs” over them, passing from one cell to the next on its way out the door. Could this be a detached bronchial cell?
A search within YouTube turned up another video, this one from the Diagnostic Bacteriology Laboratory at Singapore General Hospital. It shows a large clump of bronchial epithelium with the cilia busily working:
In this sample, the cells are all fixed in place and firmly attached to one another, so we don’t see the feeble rocking movement of the free-floating cell. Still, I’m persuaded me that the cells in both videos, as well as the one I’d seen back in 1990, are all ciliated nasal or bronchial cells.
And this brings me to the new puzzle. A comment on the second video sent me to an intriguing post at a pathology blog called Microcosm, where I learned something fascinating and, I’m sorry to say, a bit revolting. It seems there have been medical case reports of humans being infected by a lushly flagellated protozoan called Lophomonas blattarum, normally found living commensally in the intestines of cockroaches or termites.
Lophomonas blattarum, from William Saville Kent, A Manual of Infusoria, 1881.
L. blattarum has been known since 1860, when it was found and described by the great Samuel Friedrich Stein. Reports of respiratory infections in humans are more recent, dating back only as far as 1993 when the first case was reported in the Chinese Journal of Parasitology and Parasitic Diseases. Since then, many dozens of similar cases have been reported, which has lead to a growing belief, among some researchers, that Lophomonas blattarum constitutes “a potentially important cause of bronchopulmonary infection and respiratory symptoms.” (Martinez-Girón and van Woerden, 2013a).
The problem is, it might not be a genuine infection at all. In all of the published case reports of Lophomonas infection, the organisms were identified by light microscopy, either from living samples or stained preparations. To date, there have been no molecular studies of the purported parasite. Identifications have been based entirely on morphological features and symptomology.
One ultrastructural study has been done, but it does not support the theory that lophomoniasis is a genuine infective disease. The 12-author paper was published in Chinese, but from the English abstract (and by laboriously feeding the text to Google Translate one paragraph at a time), I gather that the researchers used optical and electron microscopy to examine mobile cells from BALF of 6 patients, which they found to be morphologically unlike Lophomonas. They also reviewed the existing literature on Chinese cases of lophomoniasis, and reached the following conclusion: “In the past 20 years, all the diagnosed cases as pulmonary Lophomonas blattarum infection reported in our country were misdiagnosed. Currently, there is no evidence to show Lophomonas blattarum as a pathogen resulting in pulmonary infection.” (Mu XL, et al., 2013) According to this study, the Lophomonas infections recorded in the literature were actually misidentified bronchial cells.
A typically murky micrograph of cells identified as Lophomonas blattarum in He et al, 2011. Martinez-Girón and van Woerden consider these to be misidentified bronchial epithelial cells.
Other workers, while agreeing that confusion between epithelial cells and Lophomonas has occurred, continue to maintain that many cases of infection are genuine. Martínez-Girón and van Woerden, who have published more about lophomoniasis than anyone, have responded to their Chinese colleagues by reaffirming their opinion that “the observation under light microscopy of this multiflagellated protozoon in symptomatic patients, who respond positively to antiprotozoal therapy, can reasonably be described as bronchopulmonary lophomoniasis.” (Martínez-Girón and van Woerden, 2013b)
The fine structure of Lophomonas blattarum (Beams et al, 1961, adapted from Kudo) . Note the wine-glass shaped calyx (CY), axial filament (AXF) and parabasal “collar” of tubules (PNT), all visible in the light microscope.
As someone who is usually more interested in the taxonomy of organisms than the pathologies they cause, I’m struck by the emphasis clinicians place on disease and symptoms rather than the creature itself. From a clinical point of view, the case for infection is strong: a given patient has respiratory symptoms, some apparently-flagellated cells are observed in bronchial lavage fluid, and the condition clears up with treatment by metronidazole, which is, after all, the preferred treatment for another metomonad parasite, Trichomonas vaginalis. These are all important facts that are hard to dismiss if you approach the subject from a clinical point of view. However, from a purely protistological perspective, we might note that identification by light microscopy is often unreliable, especially with such small cells; the micrographs in the published literature are usually quite poor, and may depict cells in an unnatural condition; the morphological characters used in these case studies tend to be weak, while stronger diagnostic characters, like the fine structures shown in the diagram to the right, are not mentioned; and, finally, the organism in question is a deep-branching anaerobe adapted to life in the intestines of insects of the order Blattodea, and it is a little suprising for it to flourish in a human lung. An organism’s habitat is a strong clue to its identity; this one is normally found in the airless hindgut of certain bugs, and possibly in the poop of certain bug-eating birds. Given that these flagellates, if that is what they are, were discovered in an untypical environment, a careful taxonomist might wait for more detailed morphological information, or, better yet, molecular data, before settling on a species-level identification. Where are the calyx and axial filament, which Richard Kudo managed to see in 1926? Where are the tubules of the parabasal body, which should encircle to the top of the cell like the ruffled collar of an Elizabethan gentleman? From what I’ve seen, they’re barely mentioned in the literature on lophomoniasis.
So we have a strong clinical profile combined with unimpressive taxonomic evidence, leaving us with a question that can’t be resolved until somebody succeeds in sequencing these cells. As Martinez-Girón and van Woerden conclude, “The development of a technique to culture the organism or the use of molecular techniques is required to resolve the issue.” If anybody is working on that, it will be interesting to see the results.
Tulodens. I ran across them while skimming the Wikipedia article on Protozoa, and the name didn’t ring a bell. However, the free encyclopedia assured me that there are “an estimated 30,000 protozoan species,” so it wasn’t too surprising that I’d missed these guys.
Apparently, there are at least two of them, or two kinds, and they move slowly, with the help of flagella. Or cilia, or possibly pseudopods.
A suspicious Wikipedian had already pinned a “citation needed” tag to that first sentence, so I thought I’d better corroborate the claim. A Google search turned up hundreds of references to these Tulodens. The organism was mentioned in a 24-page brochure on the most common causes of calf diarrhea. It was listed among the germs on All About Germs. On a Q&A site, somebody asked for the name of a protozoan that moves “by lashing one or more of its whiplike parts,” to which a user named 1Jazz helpfully responded: “Tulodens are a form of protozoa that move using a whip-like tail.” That’s the internet for you, always ready to help a guy with his homework.
World Renowned Parasitologist Dr. Omar Amin. [Click Image for Source]
So, are Tulodens parasitic? We already suspect that they cause calf diarrhea, but what else is known? All sources agree that they are very slow. Evidently they are also common, and possibly heterotrophic. If you like, you can read about them in a book called Fundamentals of Plant Pathology, by N. G. Ravichandra. The paperback isn’t cheap, but the Kindle edition is a very affordable CDN $7.32. Ravichandra discusses Tulodens briefly on page 254, but unfortunately does not add to what we already know. They are “slow moving.” Well, sure.
Are these Tulodens?
But it would be nice to know what they look like. A search in Google Images called up all kinds of sciencey pictures: stained mounts, petri dishes, an Asian-looking guy peering into a microscope. Google Images also threw up a pretty cool papier-mâché model of a Vorticella made by a schoolboy in Spain as a science fair project. The boy’s blog post includes another passing reference to the famously slow-moving Tulodens, and he is careful to cite Wikipedia as his source (which makes him a more conscientious scholar than “world renowned parasitologist” Dr. Amin).
Tulodens are also known to the Germans, the Portuguese and the Russians, so presumably their distribution is cosmopolitan. Curiously, all these sources seem to have pulled their information from the same place: the English Wikipedia article on Protozoa.
Not a Tuloden, but quite a nifty Vorticella. Well done, Álvaro! [Click Image for Source]
I wasn’t learning anything new, so I turned to the biodiversity databases. I checked the Catalogue of Life, the Encyclopedia of Life,AlgaebaseandWoRMS. I’m happy to say that Tulodens did not appear in any of them. Perhaps some Wikipedia editor had simply misspelled the name of some legitimate protist? I ran my eye down a few alphabetized species lists, but didn’t see any likely candidates.
At this point, I decided to see what I could learn about the Wikipedian who had introduced this organism to the world. Luckily, Wikipedia discards almost nothing: every version of every page is saved, so it is easy to retrace the entire history of an article back to the very moment it was created. The protozoa article has been edited thousands of times since it was created on May 3, 2001, but it did not take me long to find the original appearance of Tulodens, on Feb 21, 2010. It read:
Tulodens are one of the slow moving form of protozoans. They are mainly found in Leyte, Philippines. Treatment would be axing, retrenching and other form of lay away operations. Exposure usually takes two years to get rid of and are usually costly to the host.
A couple of weeks later, another editor came along and removed the scary bit about the “axing and retrenching” treatment needed to cure a Tulodens infection. Within the month, somebody else edited out the observation that Tulodens were “mainly found in Leyte, Phillipines.” Now, nothing stood between Tulodens and the claim that these little fellows “move around with whip-like tails called flagella.” There was still a certain awkwardness in the passage, though, which described the “Tulodens” (plural, presumably) as “one of the slow moving form of protozoans.” It was two years before another editor solved that problem by ingeniously changing the phrase to read “Tulodens are 2 of the slow-moving form of protozoa.” Recently, that was changed again, to read “two of the slow-moving form of protozoa.” Wikipedia’s Manual of Style stipulates that integers below nine should be spelled out as words, and Wikipedians are very careful about things like that. Somewhere along the way, somebody added a hyphen between “slow” and “moving.”
Having satisfied myself that Tulodens don’t exist outside the internet, I removed them from the original article. There’s no way to delete them from all those other sites, though, and it will be interesting to see how long they last, now that the source is gone. During their five year run on Wikipedia Tulodens managed to wriggle into hundreds of other hosts. They’re informational parasites, you could say. They may be slow, but they get around.
Sometimes, it happens automatically. Web bots and data scrapers find these tasty scraps of knowledge as they’re scuttling around the net, and simply scoop up the content and repackage it. It’s nothing remarkable, just the machinery of the internet babbling to itself. More troubling, I think, is the way this fanciful critter–dreamed up, apparently, by a bored employee at a mid-western customer services company–has found its way into so many credible-looking commercial and educational sites. Some of these sources appear, at a glance, more professional and authoritative than Wikipedia, with better web design and nicer pictures, and none of them can be edited by their readers. That last point is the important one, I think. Some might think the moral of this little story is something along the lines of “don’t trust Wikipedia,” but that isn’t it at all. Anyone can change Wikipedia for the better, if they feel like it, but there’s not much we can do about diploma mills and fake textbooks and quack doctors who are quite happy to resell the information they picked up for free, but can’t be bothered to check its veracity. I think that tells us who the real parasites are.




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}